Tel: 36 42 402 488, Fax: 36 42 402 605
This article published in: IBIS, 137: 162-168. (1995)
Summary
The aim of this study was to investigate the effects of Sahelian rainfall on the Central European Sand Martin Riparia riparia population for the period 1986-1992. The studied population breed along the River Tisza in Hungary and is one of the largest in Central Europe. The changes in the population size did not show a significant relationship to Sahelian rainfall but a large decrease in 1991 coincided with the drought in three Sahelian areas. The survival rates of adults had a significant relationship to the rainfall of the Southern Sahel but not to the other two studied areas (Northern Sahel, Central Sudan). The model selection by the SURGE program included this relationship and raised the possibility of predicting the adult survival rate in autumn for the following wintering season, using African rainfall data. The analysis showed a significant difference in the survival rates of the sexes, with that of the female being lower. Differences in the changes of the population size and adult survival rate show the importance of the recruitment of adults and this may depend mainly on the survival of juveniles.
Introduction
The large decline in Sand Martin Riparia riparia populations between 1968 and 1969 associated with drought condition in the Sahel (Kuhnen 1975, Cowley 1979) demonstrated for the first time the importance of the weather conditions in the African winter quarters on long-distance Palaearctic migrants (Moreau 1972, Berthold 1973, Winstanley et al. 1974). This relationship has now been established for other migrant populations, including the Purple Heron Ardea purpurea, Swallow Hirundo rustica , White Stork Ciconia ciconia and Sedge Warbler Acrocephalus schoenobaenus (Den Held 1981, Cavé 1983, Møller 1989, Kanyamibwa et al. 1990, Peach et al. 1991). The same relationship has subsequently been found for other Sand Martin populations in western and northern Europe thus confirming the sensitivity of this trans-Saharan migrant species (Cramp 1988) to African weather conditions (Svensson 1986, Jones 1986, Persson 1987). With the exceptions of the studies of Cowley (1979) and Persson (1987) who estimated survival rates, these studies are based on changes in population size. However, to understand the mechanisms by which African conditions affect population processes and to be able to make a short-term forecast, more detailed studies are needed. An essential part of these surveys are adult survival rates, which are one of the most important and, at the same time, the most problematic parameter used to study the influence of migration and wintering on populations of migrants (Baillie & Peach 1992, Bibby 1992). In the case of the Sand Martin, there has been a good opportunity to use capture-recapture methods to estimate both population size and survival rates (Szép 1991). Collecting high quality data and using recent methods (Lebreton et al. 1992) there is an unique opportunity of obtaining unbiased and precise annual survival rates of this migrant passerine and to analyse the effects of different environmental factors on survival.
In this paper I investigated a Sand Martin sub-population which represents about 10% of central European Sand Martin population (Tucker and Heath 1992) and is suitable for integrated monitoring work (Baillie 1990). I considered the relationship between the population size and survival rates of Sand Martins in central Europe and rainfall on the wintering grounds.
The surveyed Sand Martin sub-population breeds in natural banks along the river Tisza, the second largest river in Hungary. This sub-population is 30-50% of 60,000-80,000 pair Sand Martins which breed in Hungary (Tucker and Heath 1992). These banks are formed every spring by heavy floods. In 1990, the average colony size was 158 pairs and 67% of the population bred in colonies which were over 200 pairs. The distribution of the sub-population along the river was associated with suitable breeding sites (Szép 1991a). The survey was carried out from a motor-boat in July between 1986-1992 and the following data were collected: location of the breeding site, length and surface of the breeding site, and number of holes in the colony (Szép 1991b). The number of holes were counted by the same person and the number of breeding pairs were calculated by the proportion of occupied holes, which was about 60% in this sub-population (Szép 1990).
Two data sets of the population size were collected by counting
1)- along all of the 560 km Hungarian section of the River Tisza (Szeged-Tiszacsécse) between 1990-1992 (Fig. 1),
2)- along a 76 km stretch on the upper part of the River Tisza, called the Upper-Tisza (section B in Fig.1), between 1986-1992.
The largest colony is at Tiszatelek (48 13' N, 21 47' E), and here ringing of Sand Martins has been carried out annually since 1986, during ten days between the end of June and the beginning of July (which is the fledgling period of the first brood in most years, Szép 1990, 1991b). The birds were caught in mist-nets and the sexes was identified by the presence of a brood patch in females. The site fidelity of the birds at Tiszatelek colony was high 90% of adult and 60% of juvenile individuals which were ringed at this colony and recaptured in the following breeding season were recaptured at Tiszatelek again (Szép 1990).
The number of adults and juveniles ringed each year varied between 700-1700 individuals (Table 1 ). A higher percentage of recaught adult females indicates a larger capture probability of females at the colony compared to the males (Table 1 ). In 1987 and 1991, the birds arrived two weeks later than usual and in 1991 additional ringing was undertaken in the middle of July to catch more juveniles. The numbers of recaptured individuals which were ringed in the previous years at this colony were high (Table 1 ). I have used only the ringing data which were collected during the normal period, except for juvenile ringing data from 1991. All capture and recapture data were computerised using software developed to handle the data and to prepare data matrices for different software packages (Szép 1991b).
The Jolly-Seber model (Jolly 1965, Seber 1965) was used to estimate abundance and recruitment using capture-recapture data and computer program JOLLY (Pollock et al. 1990). The group of breeding adults which potentially could be ringed or recaptured (sampled) at the studied colony in Tiszatelek during the study years, is an open population because emigration and immigration occurred.
Survival rates and recapture probabilities were estimated by fitting a modification of the Cormack-Jolly-Seber model, using programs RELEASE (Burnham et al. 1987) and SURGE (Clobert et al. 1987, Lebreton & Clobert 1986, Pradel et al.1990). Likelihood ratio tests and Akaike's Information Criterion (AIC) were used to select the simplest model providing an adequate description of the data, and a goodness-of-fit tests were conducted on the final model (Clobert & Lebreton 1985, Clobert et al.1987, Lebreton et al. 1992).
The wintering ground of western and northern European Sand Martin populations is in the Sahel zone of West Africa (Mead 1979). There are limited data suggesting that central European populations use the western African wintering ground, as Czecho-Slovakian breeding birds have been found in spring at Lake Chad (Mead 1979). There have been only two long-distance recoveries of Sand Martins ringed in Hungary. One of these bird was ringed as a juvenile at a colony in Tiszabercel (48 09' N, 21 39' E) in July 1985 was recaptured at Eilat, Israel (29 33' N, 34 55' E) in May 1986 whilst the other was ringed as juvenile at a colony in Tiszatelek (48 13' N, 21 47' E) in July 1989 and recaptured in Malta (36 03' N, 14 14' E) in May 1991. Hungarian birds may use different migration routes, but the role of the Sahel zone in the preparation for trans-Saharian flight in spring and in wintering may be essential. Drought in the Sahel, which lies at the southern edge of the Sahara, from the Atlantic to the Red Sea, has a severe effect on Sand Martin survival rates (Cowley 1979). The Sand Martin's feeding is associated with wetland habitats both in the migrating and wintering grounds and rainfall data from the Sahel provide indirect information about the condition and abundance of these habitats (Morel & Morel 1992). I used rainfall data from three separate areas of northern tropical Africa (northern Sahel, 10 - 17 N, 20 W - 5 E; southern Sahel, 3 - 10 N, 20 W - 5 E; central Sudan, 10 - 16 N, 22 - 37 E). These areas may lie on the migration route and wintering grounds of migrating Sand Martins which breed in central Europe. The rainfall data extended from May to October, when most of the annual precipitation occurs in these regions, and were in the form of Rainfall Anomaly Indices (Barring & Hulme, in press) with 1951-1980 as the reference period. The rainfall indices were applied to the year which followed the wet period. The migrating birds arrived in that area after the wet period and potential effects of the rainfall on the breeding population may detect in the following year when the migrating birds arrive on their breeding grounds.
Change in abundance using counts
The size of the sub-population which breed on the Hungarian stretch of the River Tisza changed markedly between 1990-1992 (Fig. 1). There was a large decrease (-47%) in 1991, which was followed by an increase (+13.5%) in 1992.
The maximum numbers of Sand Martins in the upper part of the River Tisza (section B in Fig.1) was 12,312 pairs in 1989. The number of birds grew slightly between 1986-1987 and there were further increases during the 1987-1989 and 1991-1992 periods (Fig. 2). A marked decline occurred between 1989 and 1991. The numbers of breeding pairs in this area was 30-50% of the total along the Hungarian part of the River Tisza. The changes in the numbers on the upper Tisza and on the other parts of the Hungarian stretch of the River Tisza were similar between 1990-1992 (Fig. 1).
The correlation between the abundance of Sand Martins on the Upper Tisza and the three rainfall indices were not significant (rs5=0.43 for Northern Sahel; rs5=0.43 for Southern Sahel; rs5=0.67 for Central Sudan) but the marked decrease from 1990 to 1991 coincided with a very dry summer in all three African regions in 1990 (Fig. 2).
Changes in abundance using capture-recapture data
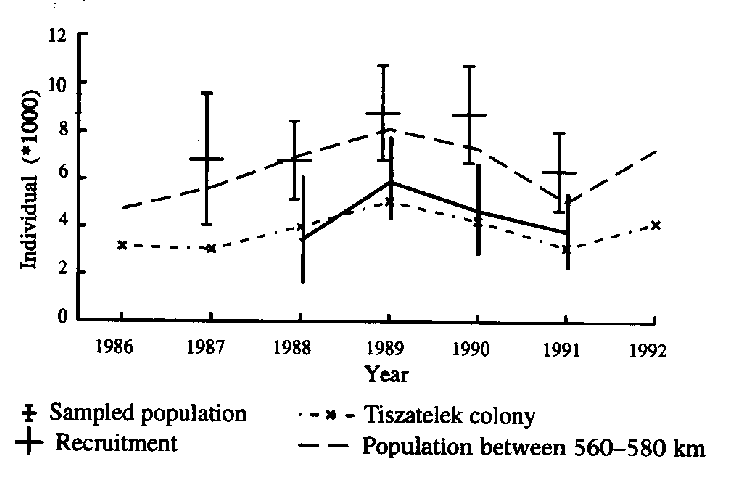
The colony size at Tiszatelek shows the same changes as found in the rest of the upper Tisza population (rs5=0.75; P<0.05) (Fig.2). Using the capture-recapture data of adult birds only, I estimated the abundance and recruitment using the program JOLLY (Model A; survival rate and recapture rate are time dependent (St,Pt); goodness of fit to the data (CHI2 df:12=16.7; n.s.). The estimated numbers shows the same changes as the surveyed numbers at Tiszatelek colony (Fig. 3). The size of the sampled sub-population is higher than the number of breeding birds at Tiszatelek colony and it is close to the number of birds which breed between the 560-580 km river stretch, including the Tiszatelek colony (Fig. 2). The highest recruitment by new breeders was in 1989 and after that there was a progressive decline until 1991. These changes coincided with the decline in the surveyed sub-population during this period. There was no detectable effect on the breeding numbers produced by the absence of second broods in 1987 and 1991. The late arrival of adults in the breeding area in 1987 and 1991 resulted in virtually no and less than 0.1% of the females raised second broods compared with the average of 20-30%.
Adult survival rates
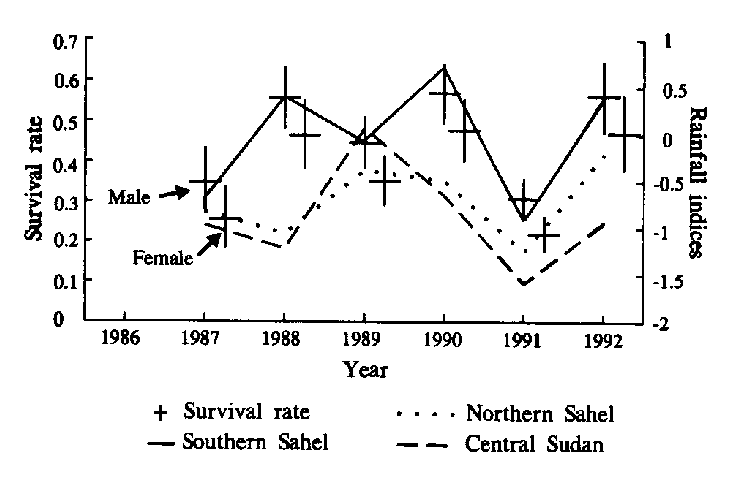
The result of TEST 3 + TEST2 from the program RELEASE (Burnham et al. 1987) suggest that the basic assumptions of the global Cormack-Jolly-Seber model (St,Pt), survival (S ) and recapture (P) rate vary with time (t), are met (CHI2 df:32=34.4; n.s.). The survival and recapture rates do not depend on the capture history of birds released on any release occasion and these parameters are specific to sampling occasions. TEST 1 from program RELEASE showed that there were no differences in survival and recapture rates between males and females (CHI2 df:11=11.4; n.s.). Because of the low sensitivity of TEST 1 to group effects in some cases, differences between males and females were investigated further by the program SURGE (Lebreton et al. 1992). I compared various capture-recapture models to analyse the time effect, sex effect, constancy, time and sex effect with interaction and without interaction on the survival and recapture rate (Lebreton et al. 1992). On the basis of Akaike's Information Criterion (AIC) the model St+s,Ps was selected which had the lowest AIC value among 25 models fitted. Using Likelihood Ratio Tests and testing model St+s,Ps against its close neighbours, no time effect in capture rates were found (CHI2 df:5=2.0, n.s.). Between the sexes, there was difference in capture rates, which was larger for females (CHI2 df:1=3.2, P=0.07). The probability of this test is above the conventional value (P<0.05), but following the recommendations of Lebreton et al. (1992), I used the P<0.1 value to assess the difference in capture rates. This difference was confirmed by the field data (Table 1 ). The survival rates of the sexes varied in parallel over time and the differences between males and females was significant (CHI2 df:1=6.28, P<0.01) (Fig.4).
In model (St+s,Ps), the survival rates are varied with time without interaction between time and sex difference between the sexes and the recapture rates are constant and different for the sexes, was thus selected and the goodness-of-fit test confirmed that this model (St+s,Ps) adequately describes the data set (CHI2 df:45=41.9, n.s.). On the basis of the St+s,Psmodel, the annual recapture rates were 0.18 [95% C.I. 0.16, 0.21] for males and 0.22 [95% C.I. 0.19, 0.26] for females during the studied years. The lowest annual survival rates were 0.31 [95% C.I. 0.26, 0.36] for males and 0.25 [95% C.I. 0.21, 0.29] for females in the 1990-1991 wintering season when the rainfall indices in the preceding wet season were the lowest in all three African regions (Fig. 4).
The estimated survival rates showed a significant positive correlation with the rainfall index of the Southern Sahel during the preceding wet season (rs4=0.94; P<0.05) (Fig. 4) but not with the other two rainfall indices (Northern Sahel rs4=0.60; n.s.; Central Sudan rs4=0.54; n.s.). After the two winters when the survival rates were the lowest (1986-87 and 1990-91) the birds returned to the breeding area two weeks later than usual and the frequency of second broods was very low.
The model selection procedure of SURGE (Table 2 ) (Clobert et al. 1987) which provide a more robust testing of the rainfall index and survival rate relation than the correlation analysis, confirmed that modeling survival as a function of rainfall in Southern Sahel with logistic linear relationship is appropriate (goodness-of-fit test,
24=1.4; n.s.) and indicate the linearity between the survival rates and rainfall index. The Akaike's Information Criterion (AIC) also supports the use of S(a+b(Rf2))+s,Ps (logistic linear relationship) model. This model had the lowest AIC value among the tested models and fit to data (CHI2 df:49=43.2; n.s.) (Table 2 ). Comparison of the selected model with nested models showed that recapture rate of females, was higher than of males (CHI2 df:1=3.39; P<0.1). On the basis of the field data there was a difference between the sexes in the recapture rates, which confirmed the result of the Likelihood Ratio Test (Table 1., 2 .). The survival rates of males were higher than those of females (CHI2 df:1=6.48,P<0.01) (Table 2 ). Leaving the difference in recapture rates out of consideration may complicate our ability to test differences in the survival rates between the two sexes (Table 2 ).
The studied sub-population along the River Tisza is an important part of the central European Sand Martin population. There was a large decrease in numbers in 1990-1991 when the population declined by a half. The same change was found in the intensively studied Upper Tisza area, which is the central part of the entire Tisza sub-population. The seven-year long study of the Upper Tisza sub-population showed similar changes to those presented in some western European countries, (e.g. Netherlands (Leys 1992), Denmark (Hustings 1992)). The population changes are not related to changes in the surface area of potential breeding sites in the Upper Tisza region which have shown continuous decline since 1987 (Szép in press). The similarity of changes in various European Sand Martin populations suggests a global effect.
The weather conditions in the African part of the Sand Martin's migration route and wintering areas may be related to the population size and survival rate (Kuhnen 1975; Cowley 1979). There was not a significant correlation between population size and weather conditions in any of three African regions, but the significant decrease in numbers in 1991 coincided with the very low rainfall in the three African regions.
The adult Sand Martin capture-recapture data from the largest colony at Tiszatelek are representative for the Upper Tisza sub-population and it is appropriate for the estimation of population size, recruitment and survival rate by capture-recapture models. The estimated abundance of the sampled population by Jolly-Seber model is nearly twice that of the breeding birds at Tiszatelek colony and shows the same changes. The sampled population may correspond to the number of breeding birds at Tiszatelek colony and nearby colonies and it may indicate the close inter-relation among these colonies. The changes in the abundance shows a close relationship with recruitment during the 1988-1991 period which reflects its importance in population changes.
The estimated adult survival rates are different for the sexes and varied the same way throughout the study. The lower survival rates of females comparing to males may reflect the effects on the condition of the females of their higher parental investment during the breeding period. The difference in the recapture rates between the sexes arises from either the different rates of nestling feeding, a larger number of males because of unpaired males, dissimilar behaviour responses to being caught, or the higher temporary emigration rate of males which decreases their recapture probability.
The adult survival rates were strongly correlated with rainfall in the Southern Sahel in the preceding wet season but not with that in the Northern Sahel and Central Sudan. Rainfall in the Southern Sahel region has a great effect on the magnitude of floods on the Rivers Senegal and Niger. These areas, with their related habitats, are some of the most important foraging and roosting places of migrating and wintering Sand Martins in western Africa (Morel & Morel 1992). The availability of non-arid habitats in Africa may have a crucial role in the success of Sand Martin foraging during the winter and the preparation for crossing the Sahara. The survival data provide indirect and circumstantial data that the Hungarian adult Sand Martins may winter in some of these areas and may use the same migration routes as the western and northern European population.
Differences in numbers and adult survival rates show the importance of recruitment and the survival of juvenile birds. The estimated population size and recruitment data suggest that recruitment of young is an important factor influencing the population size. The absence of a second brood in some years, which resulted in a lower nestling production, did not have an obvious effects on population size in the following year. These results suggest that juvenile survival may be a greater influence on population change than the number of nestling fledged. The body condition of juveniles and the environmental conditions in the breeding and migrating areas may play a crucial role in their survival, and, thus recruitment.
The relation between the dry weather in the Sahel and Sand Martin survival rate is the same as that found for the Purple Heron Ardea purpurea, White Stork Ciconia ciconia and Sedge Warbler Acrocephalus schoenobaenus (Den Held 1981, Cavé 1983, Kanyamibwa et al. 1990, Peach et al. 1991).
The results from the studied Sand Martin sub-population raises the possibility of predicting the adult survival rate during the following wintering season in autumn by African rainfall data. Forecasting a considerable decline in adult survival rates several months before the migrants arrive, could initiate actions which may help the migrants to recover their population size. Developing methods to forecast survival rates, recruitment and population size of the Sand Martin and to check these predicted parameters would be a very valuable help to studies or the protection of other Trans-Saharan migrants.
I am grateful to Edit Molnár, Zsolt Sóvári, Zoltán Barta and volunteers of the Nyíregyháza Chapter of the Hungarian Ornithological Society for their assistance in the population survey and ringing work in the Akcio Riparia camps. I would like to thank Dr. M. Hulme of The Climate Research Unit, University of East Anglia, for providing African rainfall indices, to Dr. W. Peach for information and valuable comments on the manuscript and to Dr. D.R. Anderson, Dr. J. Clobert, Dr. J.-D. Lebreton, Dr. R. Pradel for providing advises on the use of program SURGE and for helpful comments and discussion on the manuscript. I am grateful to Dr. A.P. Møller, Dr. M. Reed, Dr. J.C. Coulson and to referees for valuable comments on the manuscript and to Matthew Britschgi and Gerard Gorman for help with translation. This study is supported by a grant from the Hungarian Academy of Science, OTKA #1147.
Baillie, S. R. 1990. Integrated population monitoring of breeding birds in Britain and Ireland. Ibis, 132: 151-166.
Baillie, S. R. and Peach, W. J. 1992. Population limitation in Palaearctic-African migrant passerines. Ibis, 134 suppl.1: 120-132.
Barring, L. and Hulme, M. (in press). Filters and approximate confidence intervals for interpreting Rainfall Anomaly Indices. J. Climate Appl. Meteor.
Berthold, P. 1973. Über starken Rückgang der Dorngrasmücke Sylvia communis und anderer Singvogelarten im westlichen Europa.- J. Orn. 114: 348-360.
Bibby, C. J. 1992. Conservation of migrants on their breeding ground. Ibis, 133 suppl. 1.: 29-34.
Burnham, K. P., Anderson, D. R., White, G. C., Brownie, C. and Pollock, K. H. 1987. Design and analysis methods for fish survival experiments based on release-recapture. American Fisheries Society Monograph 5.
Cavé, A. J. 1983. Purple Heron survival and drought in tropical West Africa. Ardea, 71: 217-224.
Clobert, J., Lebreton, J. D. 1985. Dépendance de facteurs de milieu dans les estimations de taux de survie par capture-recapture. Biometrics, 41: 1031-1037.
Clobert, J., Lebreton, J. D., Allaine, D. 1987. A General Approach to Survival Rate Estimation by Recaptures or Resightings of Marked Birds.- Ardea 75: 133-142.
Cowley, E. 1979. Sand Martin population trends in Britain, 1965-1978. Bird Study, 26: 113-116.
Cramp, S. (ed) 1988. Handbook of the Birds of Europe the Middle East and North Africa. Volume 5. Oxford, Oxford University Press.
Den Held, J. J. 1981. Population changes of the Purple Heron in relation to drought in the wintering area. Ardea, 69: 185-191.
Hustings, F. 1992. European monitoring studies on breeding birds: an update. Bird Census News, 5: 1-56.
Jolly, G. M. 1965. Explicit estimates from capture-recapture data with both death and immigration-stochastic model. Biometrika 52: 225-247.
Jones, G. 1987. Selection against large size in the sand martin (Riparia riparia) during a dramatic population crash. Ibis 129: 274-280.
Kanyamibwa, S., Schierer, A., Pradel, R. & Lebreton, J. D. 1990. Changes in adult annual survival rates in a western European population of the White Stork (Ciconia ciconia). Ibis, 132: 27-35.
Kuhnen, K. 1975. Bestandsentwicklung, Verbreitung, Biotop und Siedlungsdichte der Uferschwalbe (Riparia riparia) 1966-1973 am Niederrhein. Charadrius 11: 1-24.
Lebreton, J. D., and Clobert, J. 1986. User's manual for program SURGE. Montpellier, France: Centre d'Ecologie fonctionelle et Evolutive, Centre National de la Recherche Scientifique.
Lebreton, J. D., Burnham, K. P., Clobert, J. and Anderson, D. R. 1992. Modeling survival and testing biological hypotheses using marked animals: A unified approach with case studies. Ecol.l Monogr., 62: 67-118.
Leys, H. N. 1992. De Oeverzwaluw Riparia riparia in 1991 in Netherland. Literatuuroverzicht Oeverzwaluw: 198-211. Leersum, Instituut voor bos- en natuuronderzoek.
Mead, C. J. 1979. Colony fidelity and interchange in the Sand Martin. Bird Study, 26: 99-107.
Møller, A. P. 1989. Population dynamics of a declining swallow Hirundo rustica population.- J. Anim. Ecol. 58: 1051-1063.
Moreau, R. E. 1972. The Palaearctic-African Bird Migration Systems. Academic Press, London.
Morel, G. J. and Morel, M. Y. 1992. Habitat use by Palaearctic migrant passerine birds in West Africa. Ibis, 134 suppl. 1: 83-88.
Peach, W. J., Baillie, S. R. & Underhill, L. 1991. Survival of British Sedge Warblers (Acrocephalus schoenobaenus) in relation to west African rainfall. Ibis, 133: 300-305.
Persson, C. 1987. Population processes in south-west Scanian sand martins (Riparia riparia).- J. Zool., Lond. (B) 1: 671-691.
Pollock, K. H., Nichols, J. D., Brownie, C., and Hines, J. E. 1990. Statistical inference for capture-recapture experiments. Wildlife Monographs, 107.
Pradel, R., Clobert, J. and Lebreton, J. D. 1990. Recent developments for the analysis of capture-recapture multiple data sets. The Ring, 13: 193-204.
Seber, G. A. F. 1965. A note on the multiple recapture census. Biometrika 52: 249-259.
Svensson, S. 1986. Number of pairs, timing of egg-laying and clutch size in a subalpine sand martin Riparia riparia colony, 1968-1985.- Ornis Scand. 17: 221-229.
Szép, T. 1990. Estimation of abundance and survival rate from capture - recapture data of Sand Martin (Riparia riparia) ringing. The Ring, 13: 204-214.
Szép, T. 1991a. Number and Distribution of the Hungarian Sand Martin (Riparia riparia (L.), 1758) Population Breeding along the Hungarian Reaches of the River Tisza. Aquila, 98: 111-124.
Szép, T. 1991b. Monitoring of abundance and survival rate of Sand Martin (Riparia riparia) population in the upper reaches of the River Tisza, 1986-1990. Ornis Hungarica, 1: 37-44.
Szép, T. (in press). Monitoring the Size, Distribution and Survival of the Sand Martin (Riparia riparia) Breeding Population in Hungary. Bird Numbers 1992. Proc. XIIth International Conference on Bird Census and Atlas Work. Noordwijkerhout
Tucker, G. M. and Heath, M. F. 1992. The conservation status of European birds, working report. ICBP and EOAC, Cambridge.
Winstanley, D., Spencer, R. & Williamson, K. 1974. Where have all the whitethroats gone ?. Bird Study 21: 1-14.
Table 1. Number of ringed individuals, percentage of individuals which were recaught at least twice in the same season (%RC), and number of recaptured individuals which were ringed in the previous years at the Tiszatelek colony.
Year No. Ringed as adult No. Ringed No. Recaptured
male (%RC) female (%RC) as juvenile
1986 361 (23) 327 (34) 928 -
1987 606 (29) 627 (39) 191 63
1988 732 (24) 722 (33) 997 167
1989 849 (28) 802 (26) 1142 211
1990 937 (24) 1068 (38) 974 326
1991 637 (36) 578 (46) 378 205
1992 962 (33) 970 (37) 1207 223
A similar number of adult female and male individuals were caught in each year (Wilcoxon matched-pairs signed-rank test (two-tailed); Z=-0.5; n=7; n.s.), but there was a difference between the sexes in the percentage of individuals which were caught at least twice (%RC) at the colony in the same season (Wilcoxon test; Z=-2.2; n=7; P<0.05).
Table 2. Summary of the information used in model selection for the capture-recapture data of Sand Martins for testing the rainfall effect on the survival rate. For each model I give the number of estimable parameters (np), the deviance (DEV), and the Akaike Information Criterion (AIC). The result of the comparison between the models is given by the 2 value (LRT) and degree of freedom (df) of the Likelihood Ratio Test. (Rf1: rainfall index in Northern Sahel; Rf2: rainfall index in Southern Sahel; Rf3: rainfall index in Central Sudan) (1 indicates P <0.1 significance level)
Model np DEV AIC LRT df Result
(1) St+s,Ps 9 7261.88 7279.88
Modelling survival as a function of African rainfall (Rfx) (logistic linear relationship)
(2) S(a+b(Rf1))+s,Ps 5 7299.79 7399.79 (2)-(1)=37.91 4 rejected
(3) S(a+b(Rf2))+s,Ps 5 7263.31 7273.31 (3)-(1)=1.43 4 accepted
(4) S(a+b(Rf3))+s,Ps 5 7309.01 7319.01 (4)-(1)=47.13 4 rejected
Modelling differences between sexes in capture and survival rates
(5) S(a+b(Rf2))+s,P 4 7266.70 7274.70 (5)-(3)=3.39 1 rejected1
(6) S(a+b(Rf2)),Ps 4 7269.79 7277.79 (6)-(3)=6.48 1 rejected
(7) S(a+b(Rf2)),P 3 7269.96 7275.96 (7)-(3)=6.65 2 rejected
Figure
Fig. 1 Population size (pairs) of Sand Martins on the different sections on the 560 km long Hungarian stretch of the River Tisza between 1990-1992. (Starting and ending points (km) of each sections are in brackets and indicated in the histogram by patterns. Section B is the upper River Tisza study area.)
Fig. 2 Population size (pairs) of Sand Martins at Tiszatelek (580 km) and at other sections of the upper River Tisza study area (544-620 km) between 1986-1992, and the rainfall indices from the preceding wet period in three different areas in Africa.
Fig. 3 Estimated abundance of the sampled population and recruitment of breeding birds (individuals) with 95% confidence limit calculated by JOLLY (Model A), number of breeding adults (individuals) determined by direct survey of breeding pairs at the Tiszatelek colony and along the 560-580 km stretch (including the Tiszatelek colony) between 1986-1992. Abundance and recruitment were calculated only for 5 and 4 years because of the limitation of the Jolly-Seber methods (Jolly 1965, Seber 1965).
Fig. 4 Estimated survival rate (previous 12 months to given year) for adult male and female Sand Martins with 95% confidence limit between 1986-1992 and the rainfall indices from the preceding wet period in three different areas in Africa.
03/09/1996
{kind=link}
{kind=link}
{kind=link}
{kind=link}